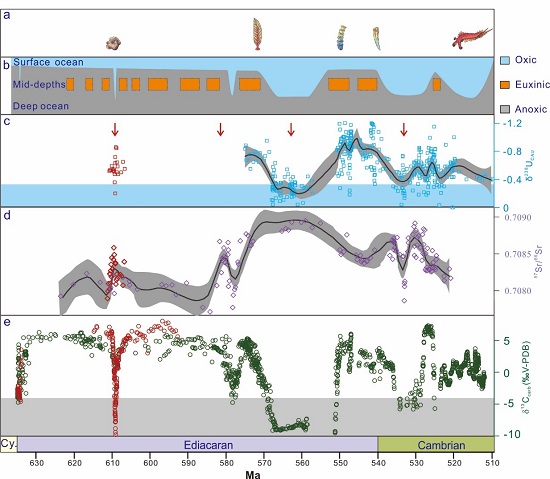
A short-lived oxidation event during the early Ediacaran and delayed oxygenation of the Proterozoic ocean. (Earth and Planetary Science Letters. https://doi.org/10.1016/j.epsl.2021.117274)
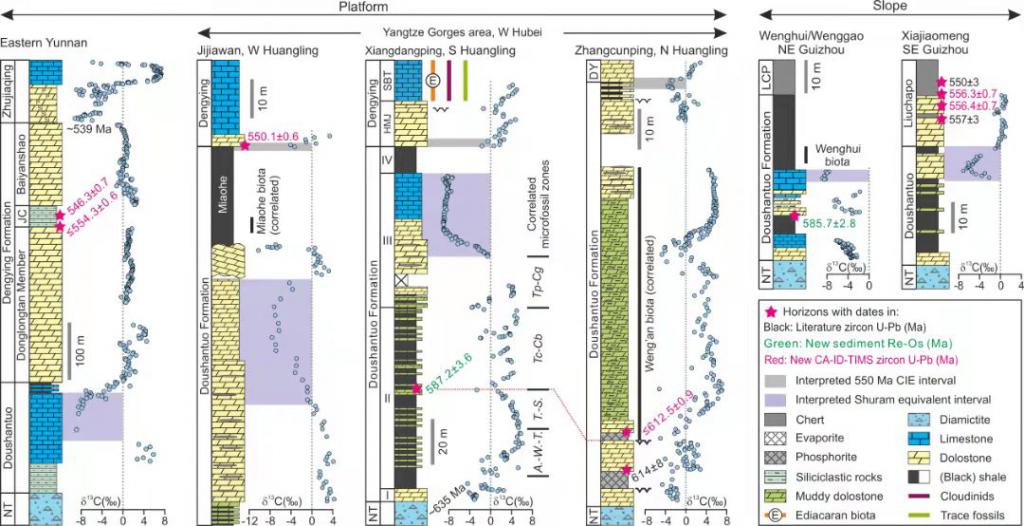
The tempo of Ediacaran evolution. (Science Advances. https://www.science.org/doi/10.1126/sciadv.abi9643)
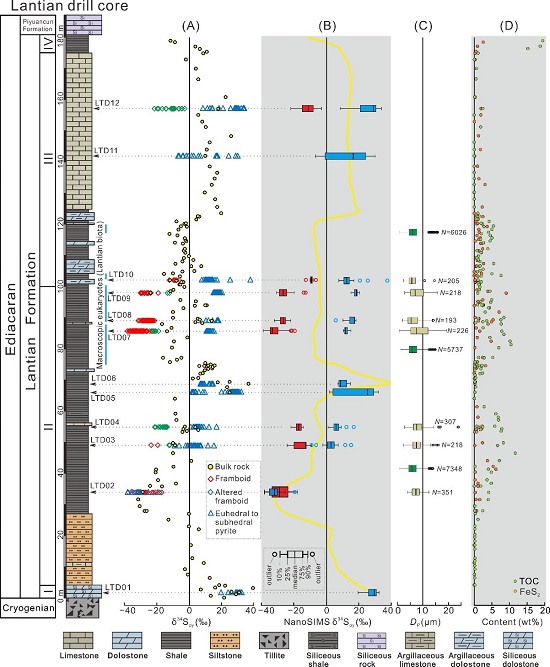
Revisiting Ediacaran sulfur isotope chemostratigraphy with in situ nanoSIMS analysis of sedimentary pyrite. (Geology. https://doi.org/10.1130/G48262.1)
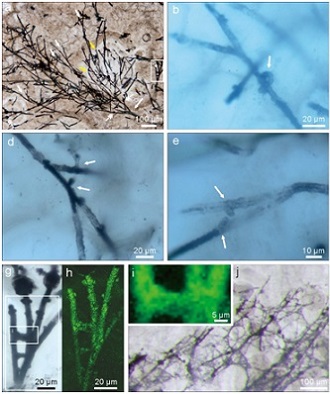
Cryptic terrestrial fungus-like fossils in the early Ediacaran Period. (Nature Communications. https://doi.org/10.1038/s41467-021-20975-1)

New Ediacara-type fossils and late Ediacaran stratigraphy from the northern Qaidam Basin (China): palaeogeographic implications. (Geology. https://doi.org/10.1130/G48842.1)
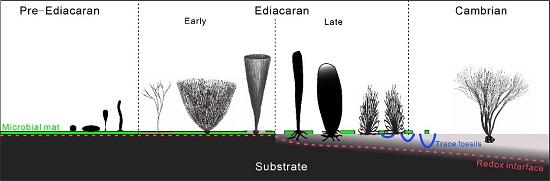
Evolution of Holdfast Diversity and Attachment Strategies of Ediacaran Benthic Macroalgae. (Front. Earth Sci. http://doi.org/10.3389/feart.2021.783427)
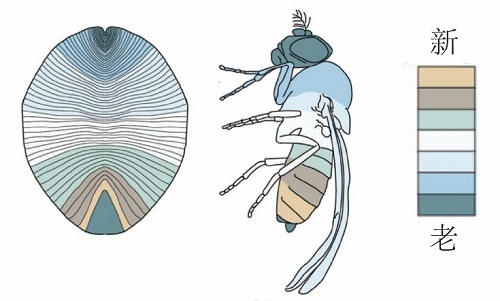
Dickinsonia from the Ediacaran Dengying Formation in the Yangtze Gorges area, South China. (Palaeoworld. https://doi.org/10.1016/j.palwor.2021.01.002)
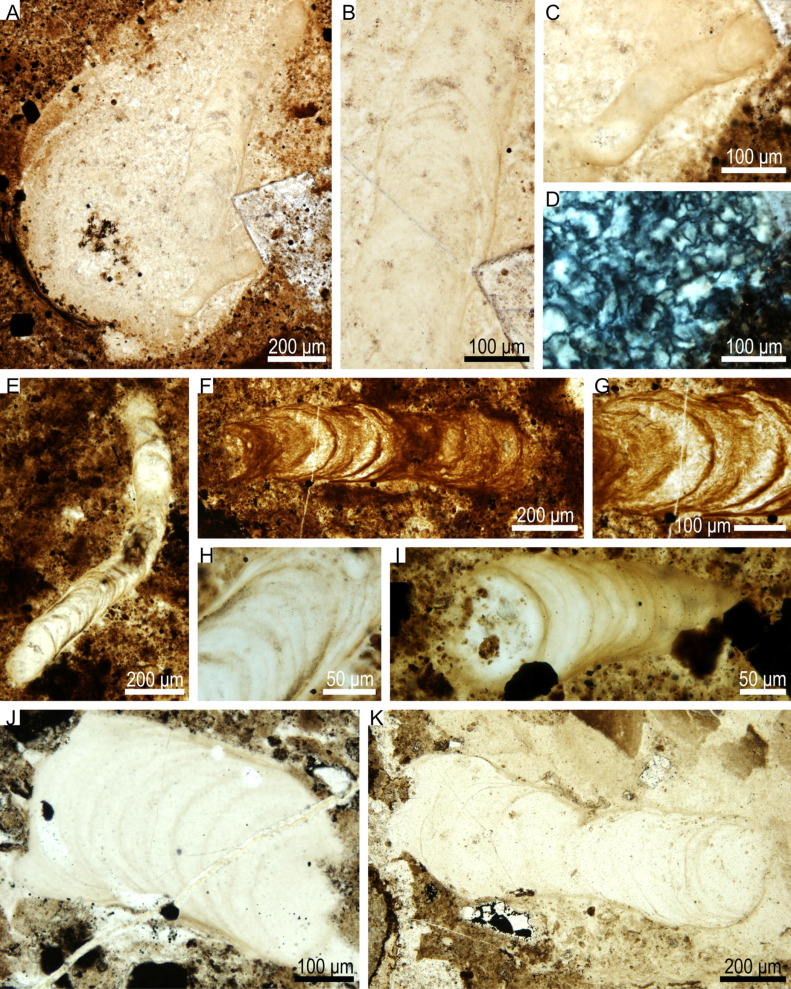
Silicified Polybessurus from the Ediacaran Doushantuo Formation records microbial activities within marine sediments. (Palaeoworld. https://doi.org/10.1016/j.palwor.2021.03.001)
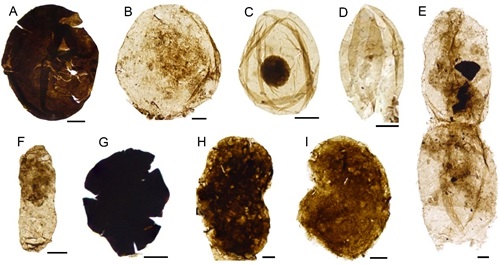
A diverse organic-walled microfossil assemblage from the Mesoproterozoic Xiamaling Formation, North China. (Precambrian Research. https://doi.org/10.1016/j.precamres.2021.106235)

A strongly positive sulphur isotopic shift in late Ediacaran-early Cambrian seawater: evidence from evaporites in the Salt Range Formation, northern Pakistan. (Geological Quarterly. http://dx.doi.org/10.7306/gq.1598)
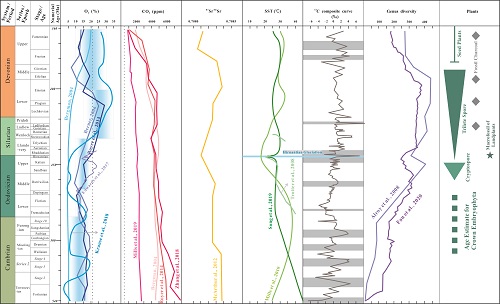
Marine oxygenation, deoxygenation, and life during the Early Palaeozoic: An overview (Palaeogeography, Palaeoclimatology, Palaeoecology. https://doi.org/10.1016/j.palaeo.2021.110715)
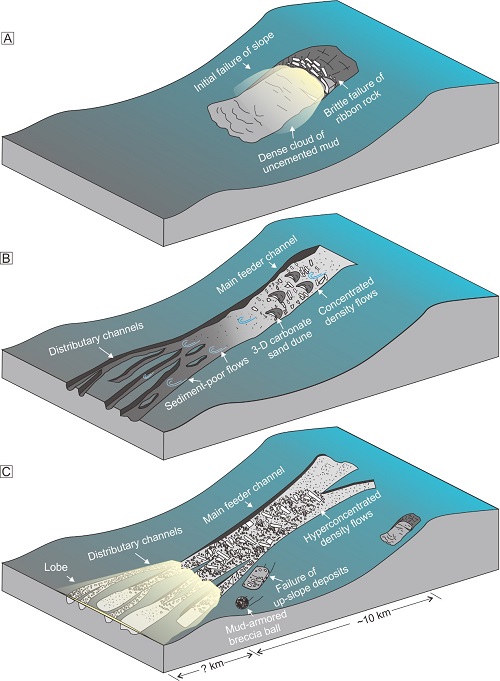
Middle Ordovician mass-transport deposits from western Inner Mongolia, China: Mechanisms and implications for basin evolution. (Sedimentology. https://doi.org/10.1111/sed.12949)
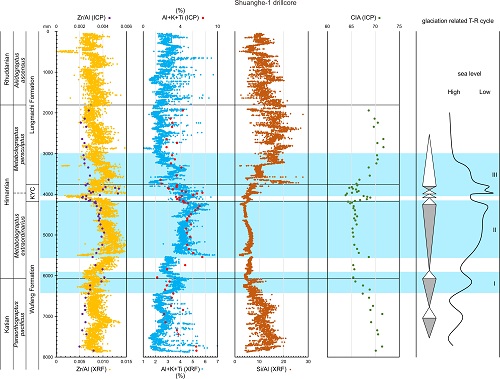
Multiple glacio-eustatic cycles and associated environmental changes through the Hirnantian (Late Ordovician) in South China. (Global and Planetary Change. https://doi.org/10.1016/j.gloplacha.2021.103668)
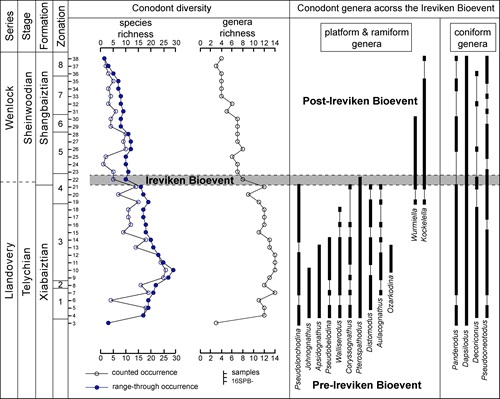
Conodont biostratigraphy and biodiversity of the middle to Upper Ordovician near Shitai of Anhui Province, South China. (Alcheringa. https://doi.org/10.1080/03115518.2020.1861325)
Middle to Late Ordovician carbon isotope chemostratigraphy of the Lower Yangtze Platform: implications for global correlation. (Geological Journal.)https://doi.org/10.1002/gj.4068)
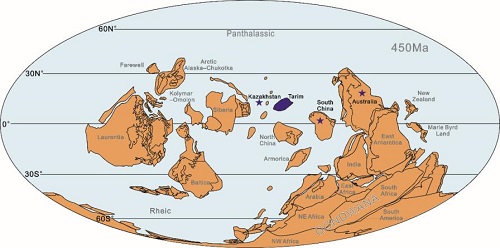
Revision of late Katian (Late Ordovician) heliolitine corals from northern Kuruktag in northeastern Tarim Basin of China. (Alcheringa. https://doi.org/10.1080/03115518.2021.1934898)
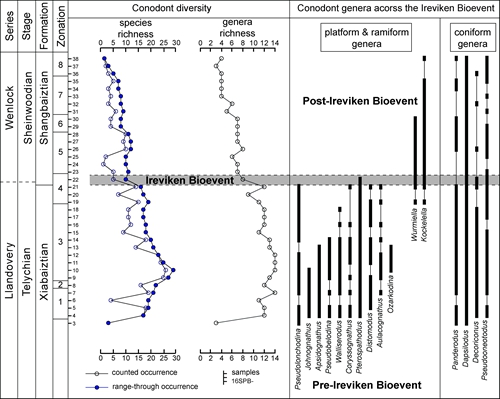
Silurian (late Llandovery – Wenlock) conodont fauna and biostratigraphy from the Yanbian area of Sichuan Province, south‐west China. (Papers in Palaeontology. https://doi.org/10.1002/spp2.1364)

Morphology and Nomenclature of Barsassia (Lycopsida) from the Middle Devonian of West Junggar, Xinjiang, China. (Plants. https://doi.org/10.3390/plants10122631)
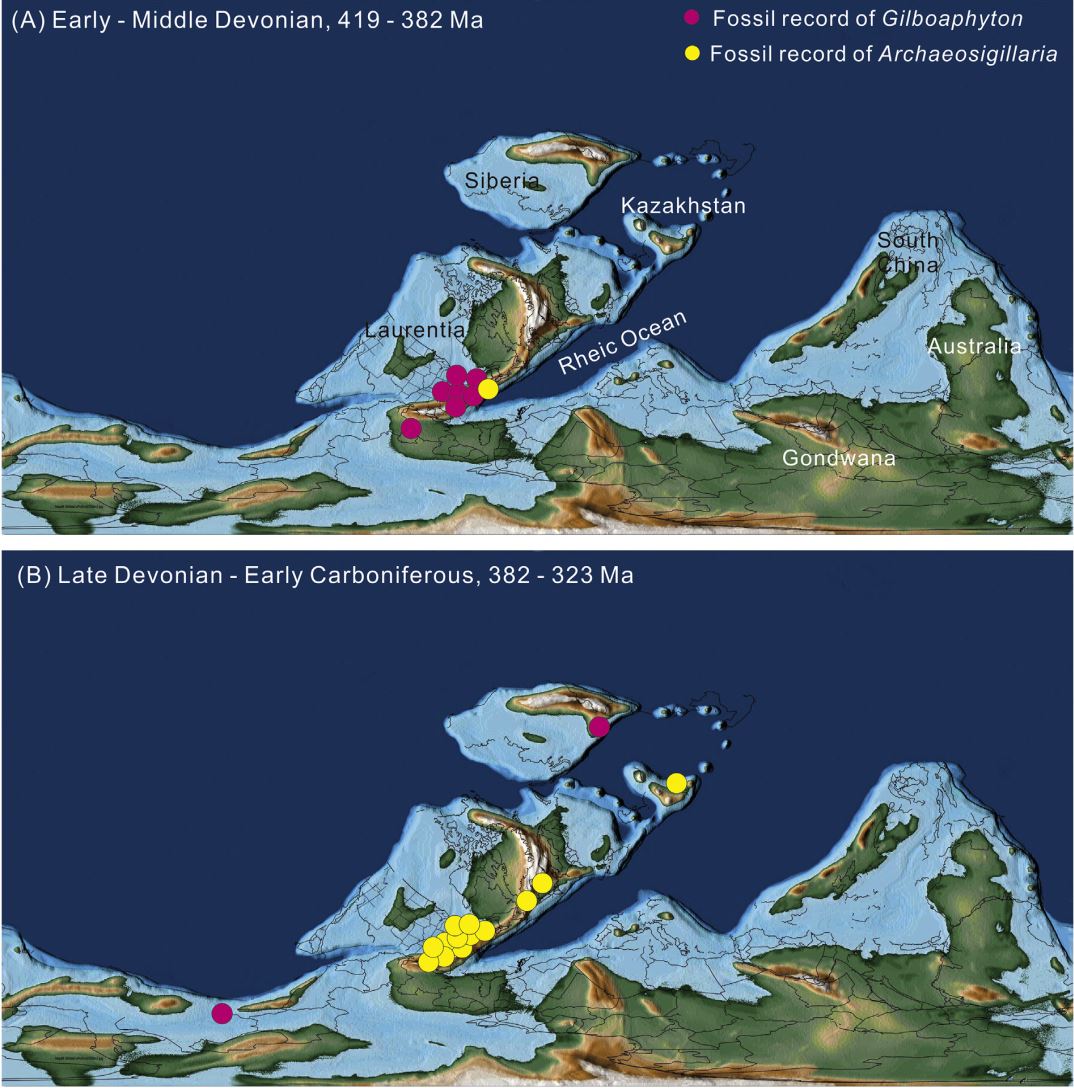
On the discovery of Gilboaphyton (Lycopsida) from the Upper Devonian of East Junggar, Xinjiang, and its global distribution. (Review of Palaeobotany and Palynology. https://doi.org/10.1016/j.revpalbo.2021.104473)
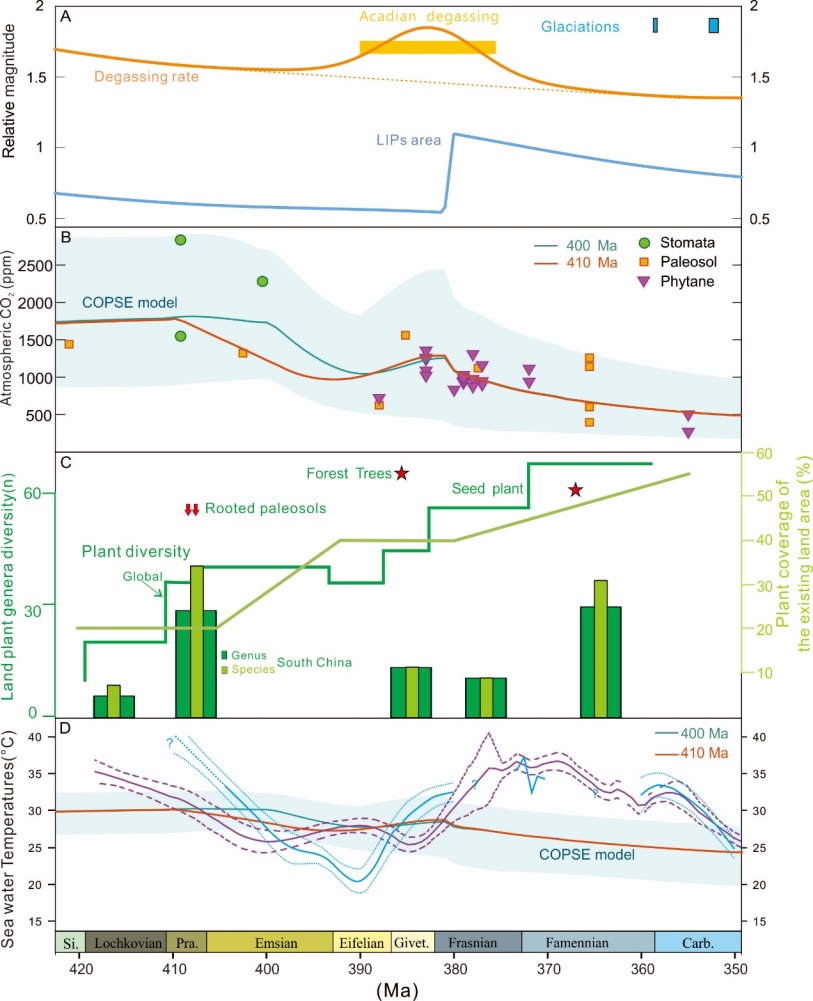
Devonian palaeoclimate and its drivers: A reassessment based on a new conodont δ18O record from South China. (Earth-Science Reviews. https://doi.org/10.1016/j.earscirev.2021.103814)

Devonian-Carboniferous boundary in China. (Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-021-00494-z)

A continuous record of the SPICE event, sea level change and the first appearance of Fenghuangella laevis in Shandong Province, North China. (Lethaia. https://onlinelibrary.wiley.com/doi/10.1111/let.12425)
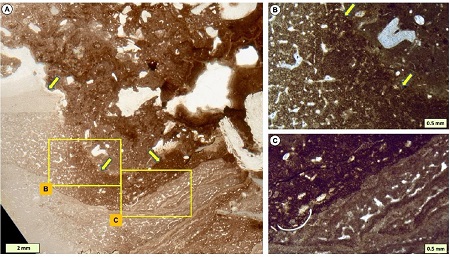
Addressing a Phanerozoic carbonate facies conundrum—sponges or clotted micrite? Evidence from early Silurian reefs, South China Block. (Sedimentary Record. https://doi.org/10.2110/sedred.2021.1.03)
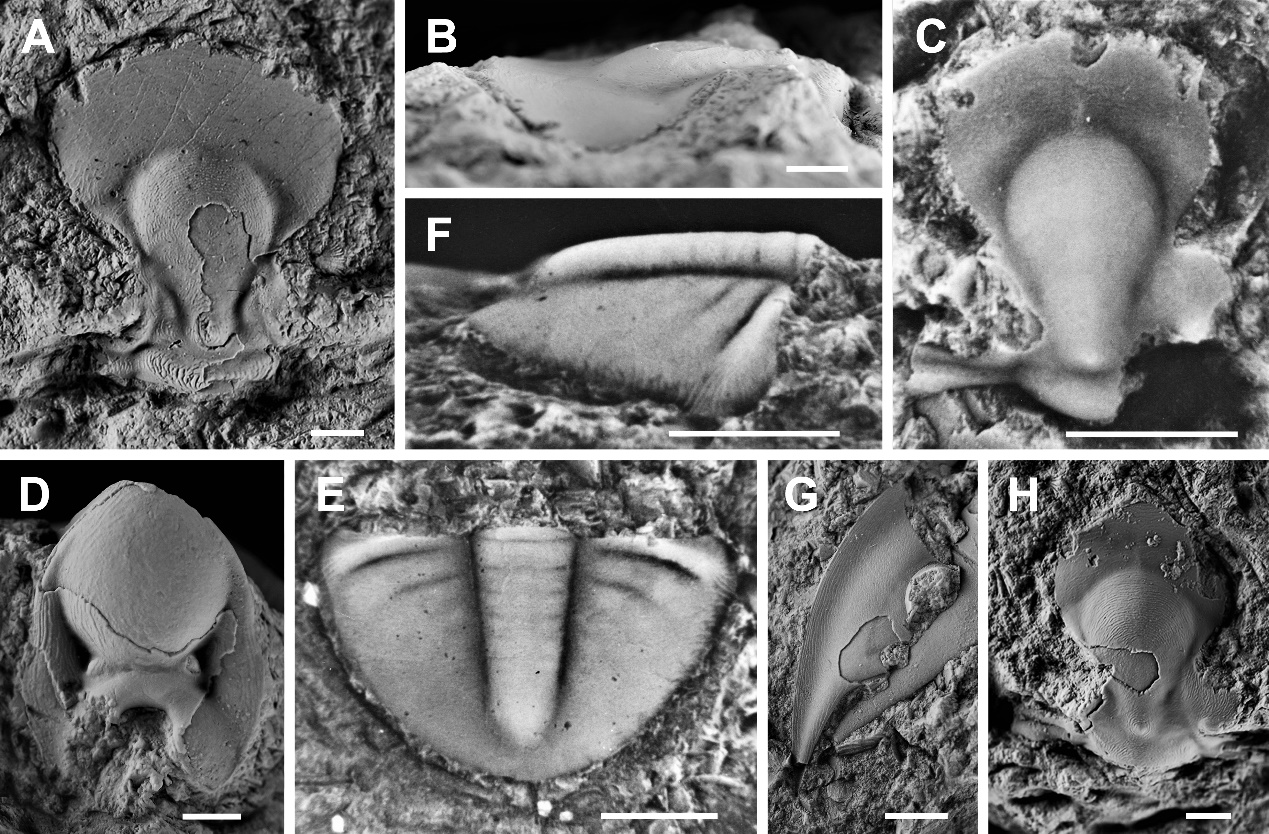
Middle Ordovician trilobites from the Houping Formation of South China: Implications for palaeoecology and palaeobiogeography. (Historical Biology. https://doi.org/10.1080/08912963.2021.1945592)
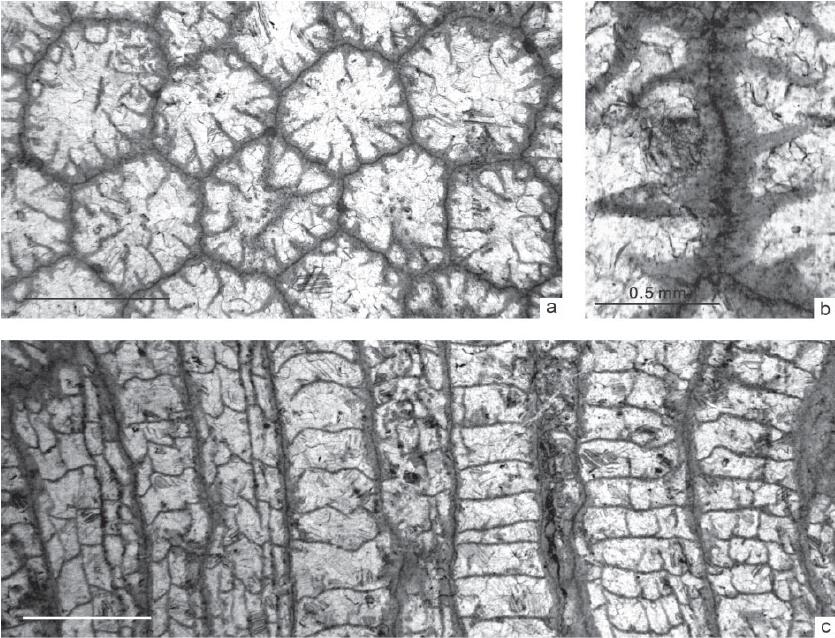
Late Ordovician corals from allochthonous clasts in the Devonian Drik-Drik Formation of northeastern New South Wales, Australia. (Proceedings of the Linnean Society of New South Wales.https://openjournals.library.sydney.edu.au/index.php/LIN/index)
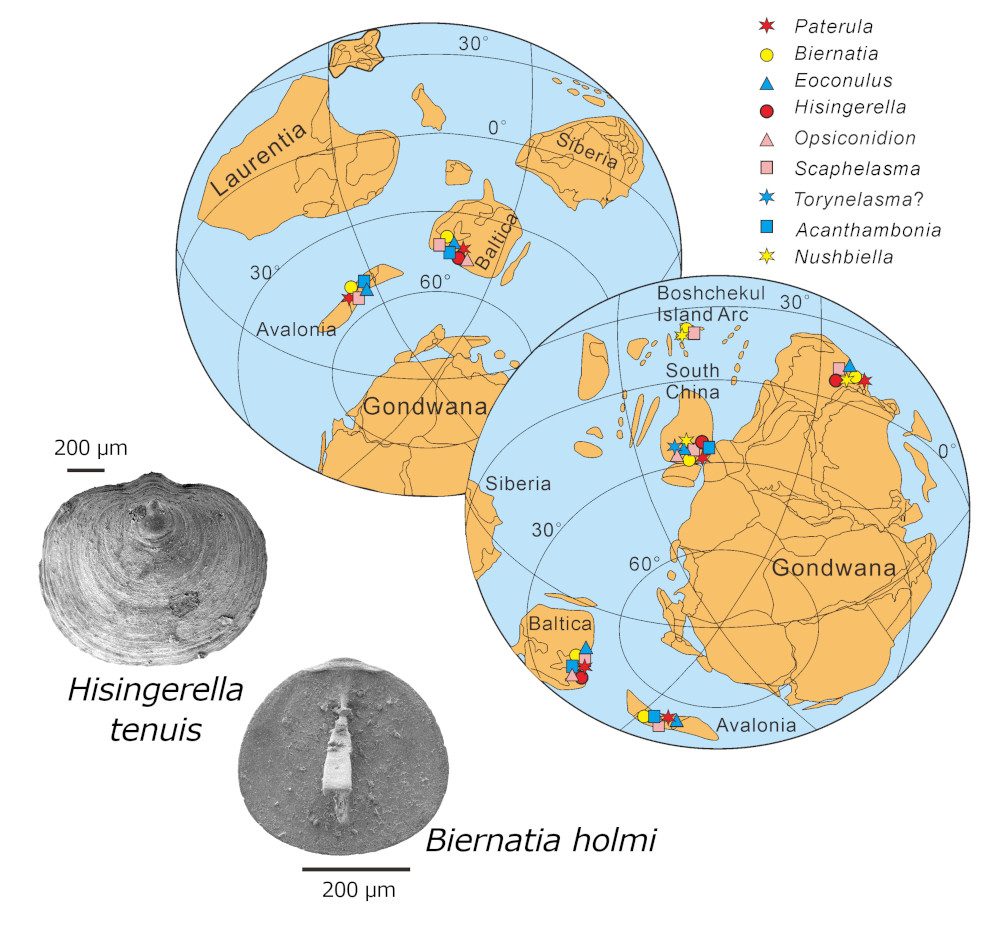
Late Ordovician (Katian) linguliform microbrachiopods from north-eastern Yunnan, South China. (Papers in Palaeontology. https://doi.org/10.1002/spp2.1407)

Phylogeny of Middle–Late Ordovician lituitid cephalopods based on cladistic analysis. (Journal of Systematic Palaeontology. https://doi.org/10.1080/14772019.2021.1944354)
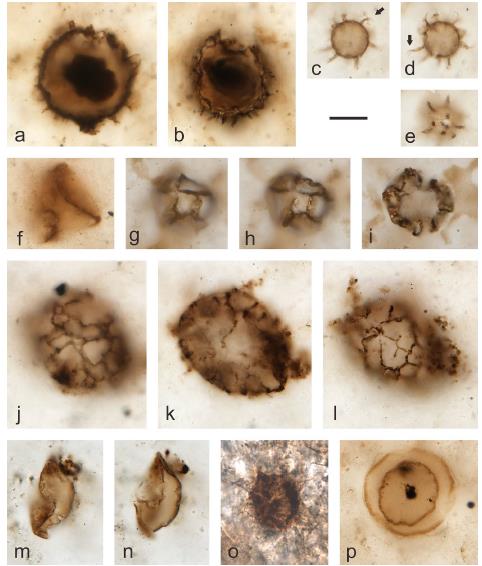
Microfossils from nodular cherts in the Middle Ordovician Yijianfang Formation carbonates (Well YJ1X), Tarim Basin, China. (Carbonates Evaporites. https://doi.org/10.1007/s13146-021-00689-7)

First mixopterid eurypterids (Arthropoda: Chelicerata) from the Lower Silurian of South China. (Science Bulletin. https://doi.org/10.1016/j.scib.2021.07.019)
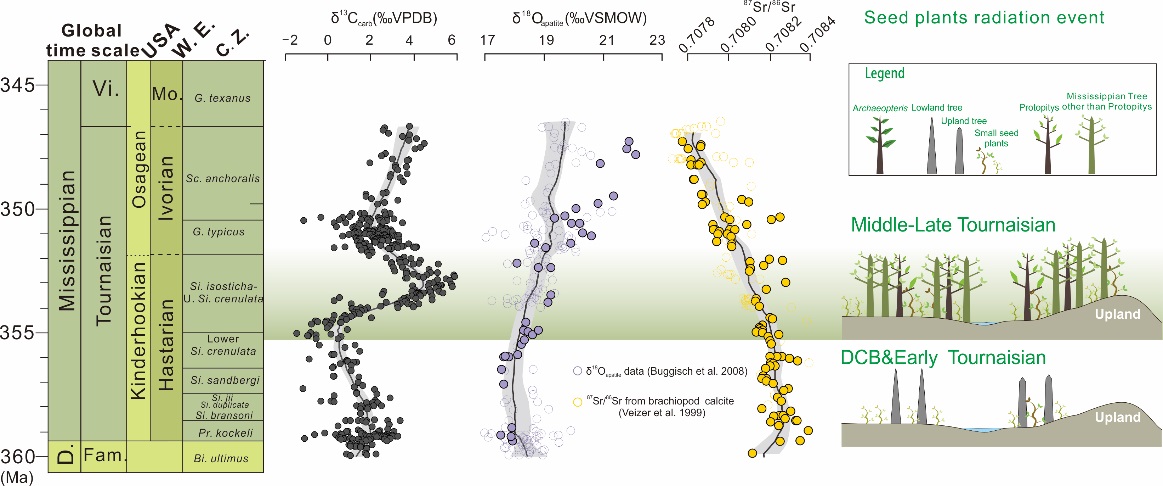
Was climatic cooling during the earliest Carboniferous driven by expansion of seed plants? (Earth and Planetary Science Letters. https://doi.org/10.1016/j.epsl.2021.116953)
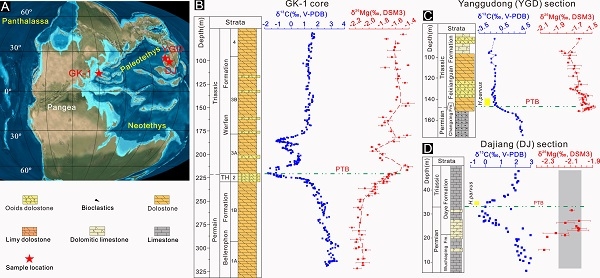
Mg isotope evidence for restriction events within the Palaeotethys ocean around the Permian-Triassic transition. (Earth and Planetary Science Letters. https://doi.org/10.1016/j.epsl.2020.116704)
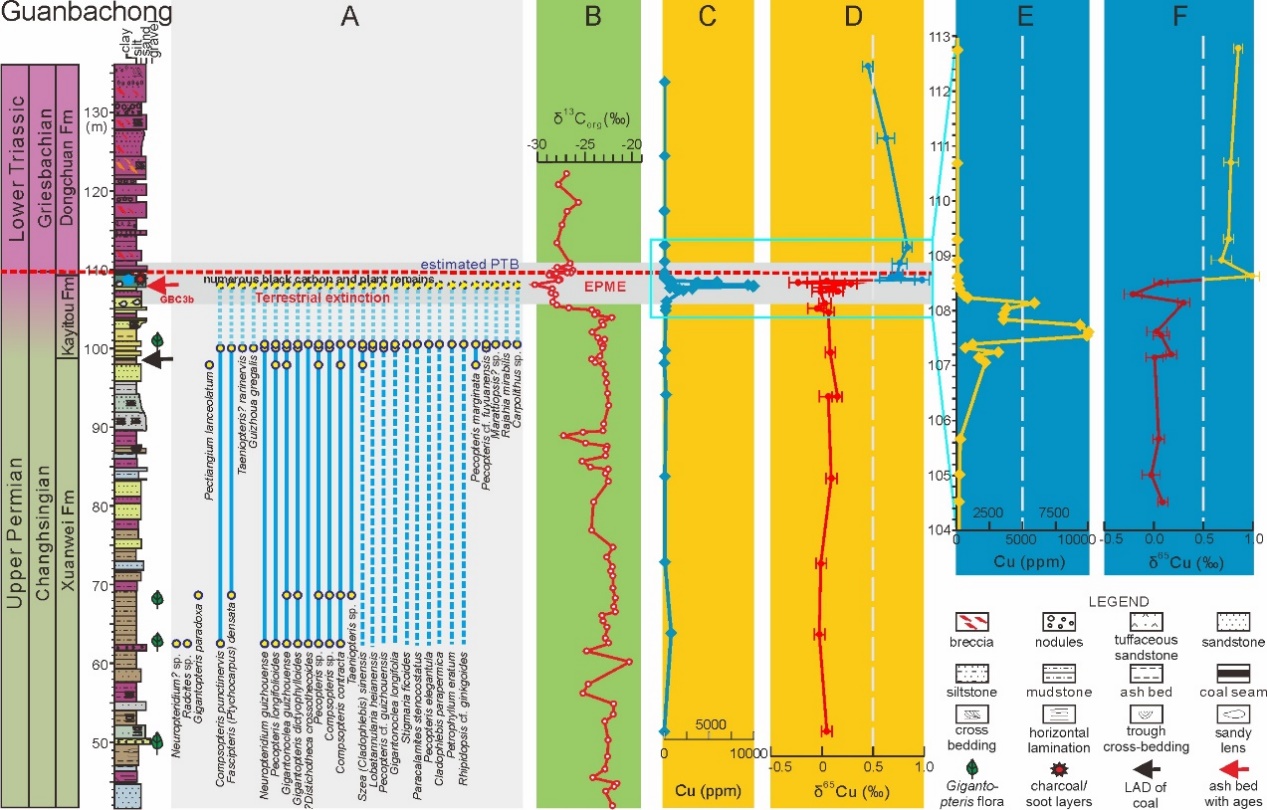
Felsic volcanism as a factor driving the end-Permian mass extinction. (Science Advances. https://science.org/doi/10.1126/sciadv.abh1390)
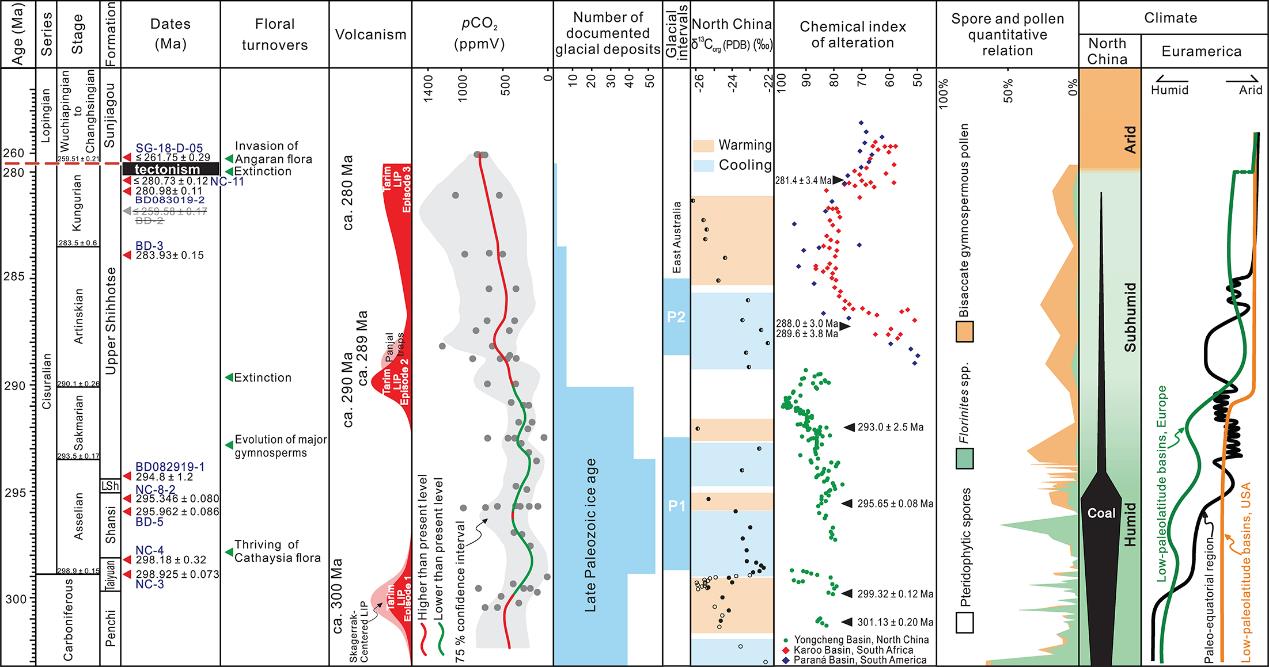
High-precision U-Pb age constraints on the Permian floral turnovers, palaeoclimate change and tectonics of North China. (Geology. https://doi.org/10.1130/G48051.1)
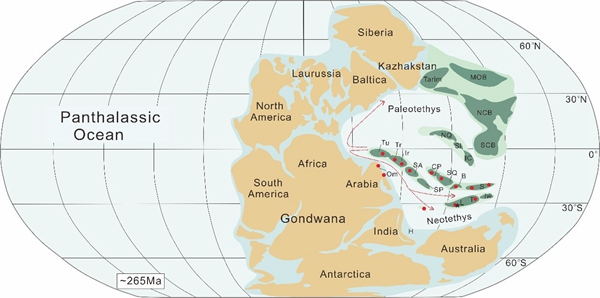
A Middle Permian assemblage of smaller foraminifera (Shanita-Hemigordiopsis assemblage) from the central Lhasa Block and its paleobiogeographic implications. (Palaeogeography, Palaeoclimatology, Palaeoecology. https://doi.org/10.1016/j.palaeo.2021.110417)
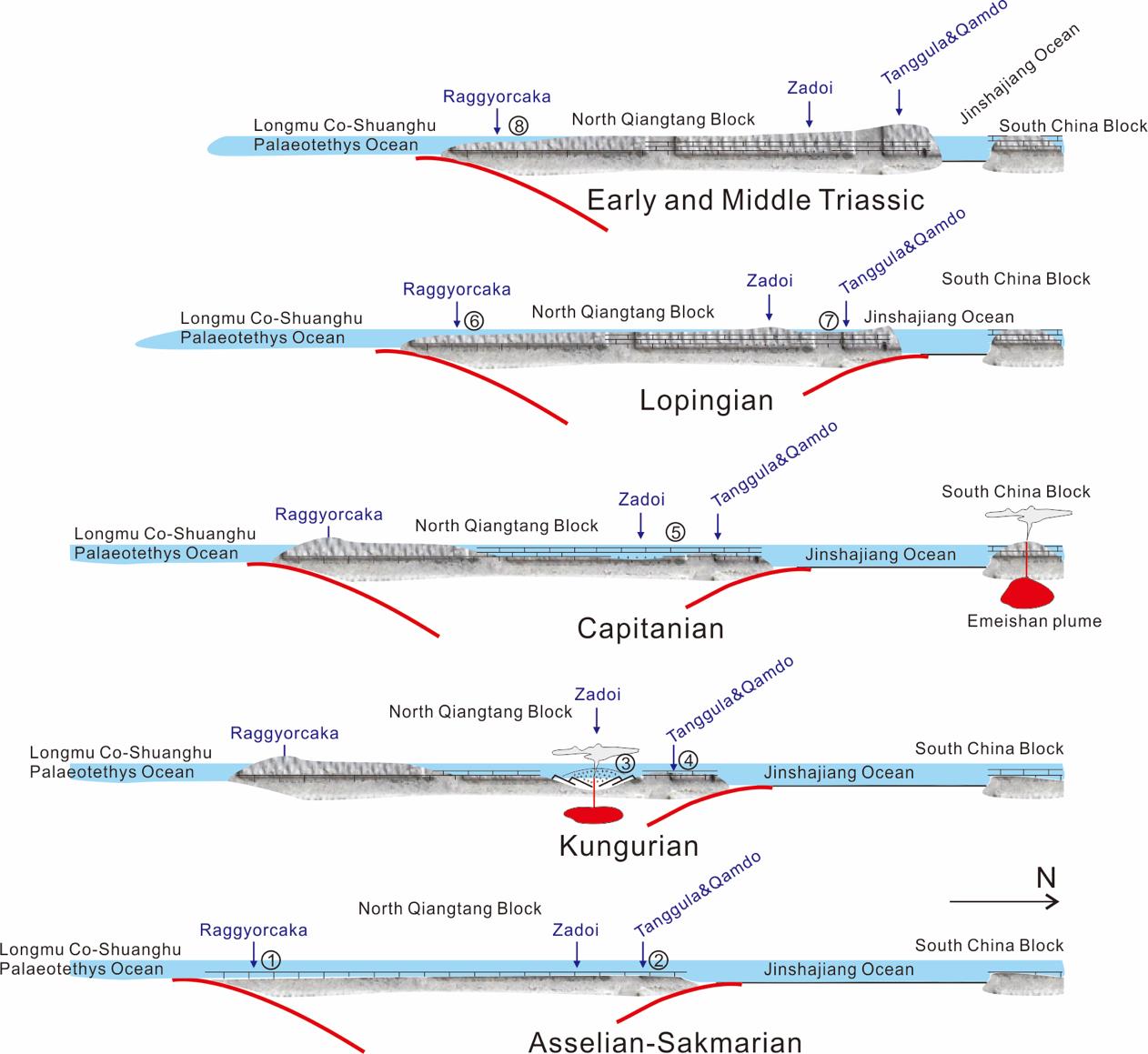
An updated age of Permian strata in the Raggyorcaka and Qamdo areas, Tibet and their palaeogeographic implications. (Palaeogeography, Palaeoclimatology, Palaeoecology. https://doi.org/10.1016/j.palaeo.2021.110660)

Ancient noeggerathialean reveals the seed plant sister group diversified alongside the primary seed plant radiation, (PNAS. https://doi.org/10.1073/pnas.2013442118)

Wuda Tuff Flora: A Permian peat-forming T0 fossil plant assemblage from Wuda Coalfield, Inner Mongolia. (Review of Palaeobotany and Palynology. https://www.sciencedirect.com/journal/review-of-palaeobotany-and-palynology/vol/294/suppl/C)

Early evolution of beetles regulated by the end-Permian deforestation. (eLife. https://elifesciences.org/articles/72692)

Wildfires and deforestation during the Permian–Triassic transition in the southern Junggar Basin, Northwest China. (Earth-Science Reviews. https://doi.org/10.1016/j.earscirev.2021.103670)
Intensive wildfire associated with volcanism promoted the vegetation changeover in Southwest China during the Permian–Triassic transition. (Frontiers in Earth Science. https://doi.org/10.3389/feart.2021.615841)
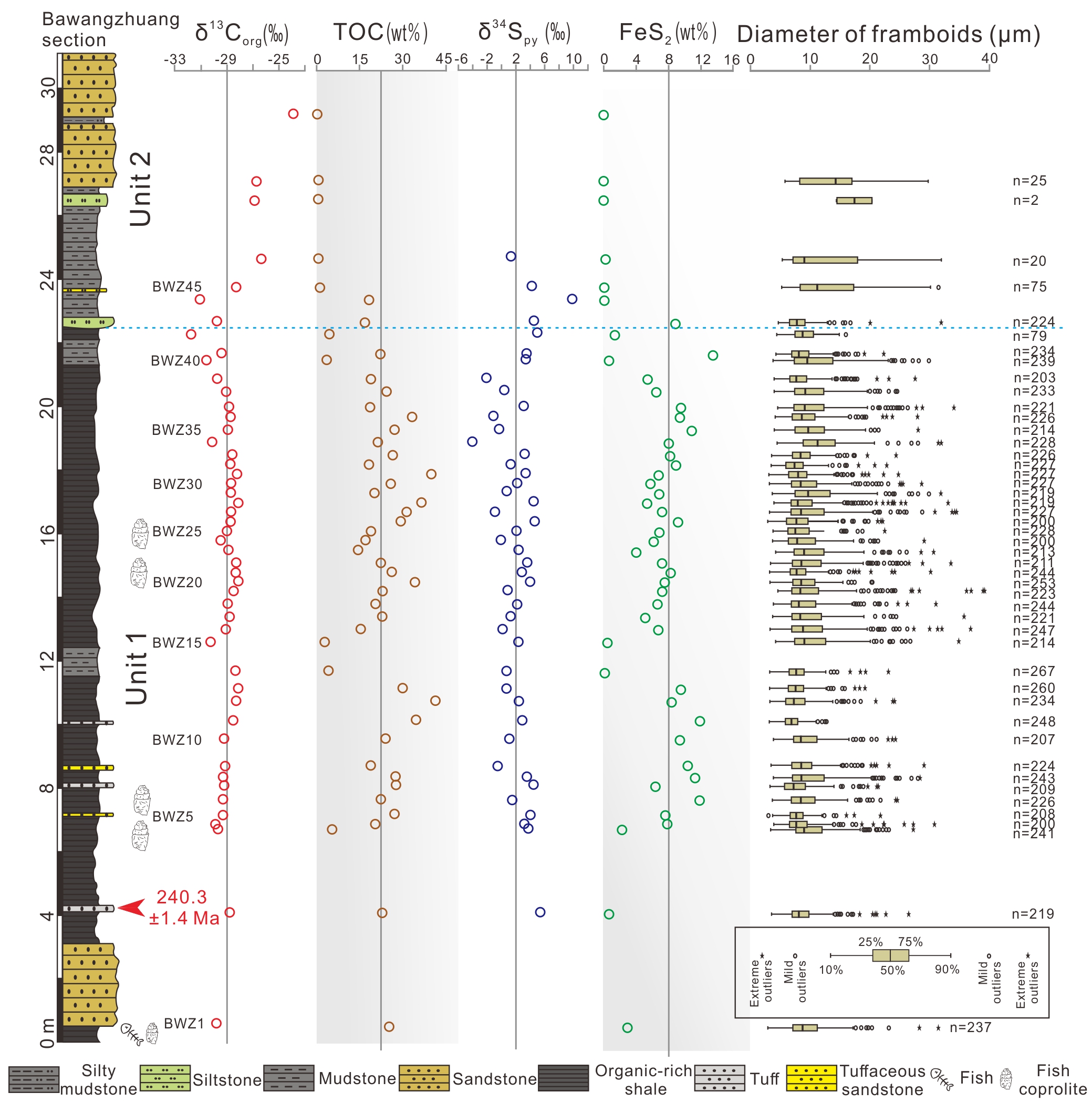
Depositional environment of Middle Triassic organic-rich shales in the Ordos Basin, Northwest China. (Geological Journal. https://doi.org/10.1002/gj.4215)
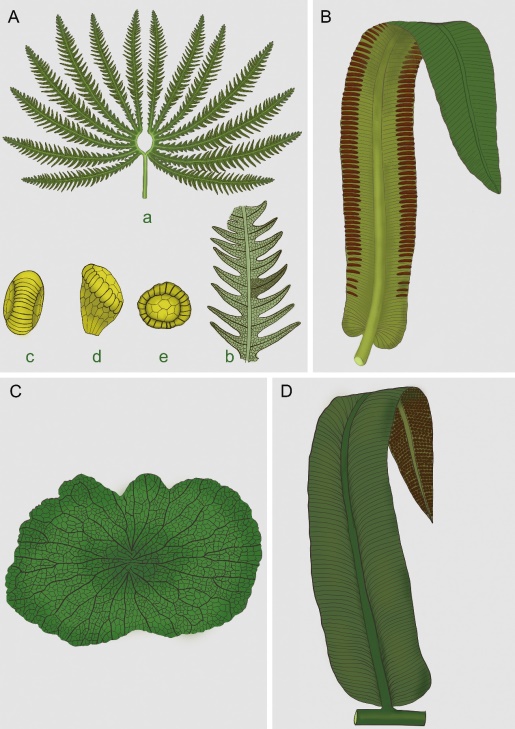
Pattern of vegetation turnover during the end-Triassic mass extinction: Trends of fern communities from South China with global context. (Global and Planetary Change. https://doi.org/10.1016/j.gloplacha.2021.103585)

Re–appraisal of Anthrophyopsis (Gymnospermae): New material from China and global fossil records. (Review of Palaeobotany and Palynology. https://doi.org/10.1016/j.revpalbo.2021.104475)

A new permineralized osmundaceous rhizome with fungal remains from the Jurassic of western Liaoning, NE China. (Review of Palaeobotany and Palynology. https://doi.org/10.1016/j.revpalbo.2021.104414)

A whole-plant monocot from the Lower Cretaceous. (Palaeoworld. https://doi.org/10.1016/j.palwor.2020.03.008)
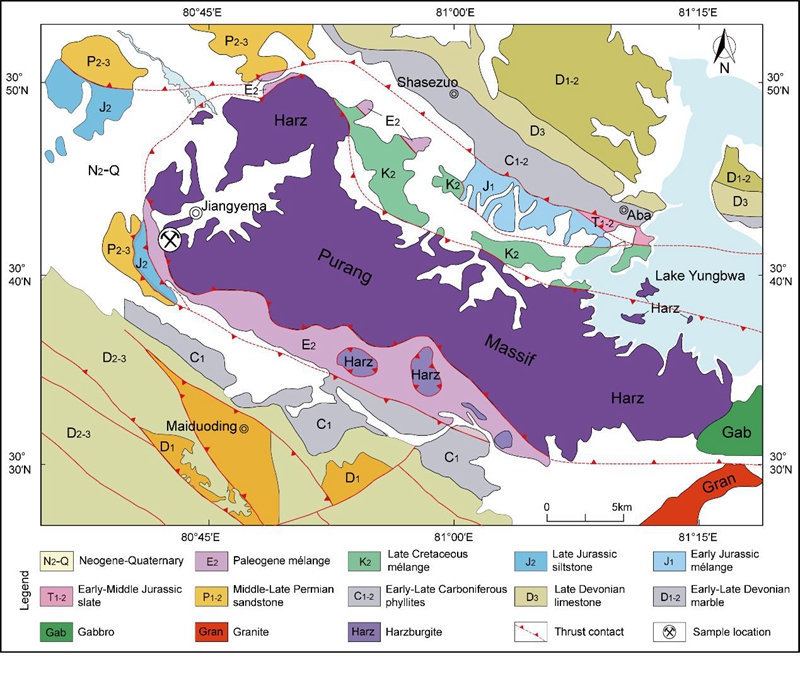
Early Cretaceous radiolarians and chert geochemistry from western Yarlung Tsangpo suture zone in Jiangyema section, Purang County, SW Xizang. (Cretaceous Research. https://doi.org/10.1016/j.cretres.2021.104840)

A latest Cretaceous gastropod fauna from the Jiaolai Basin of East Asia. (Cretaceous Research. https://doi.org/10.1016/j.cretres.2020.104736)
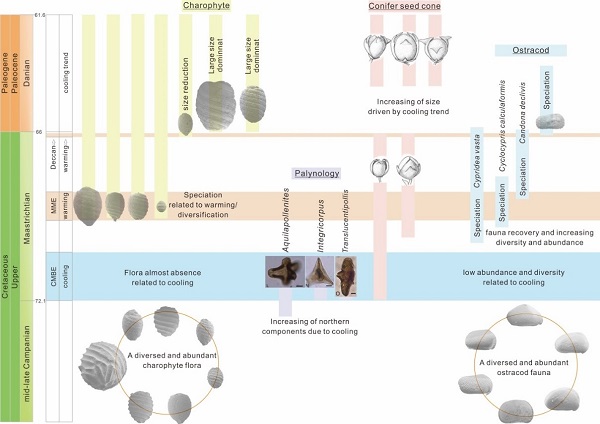
Response of the lacustrine flora in East Asia to global climate changes across the K/Pg boundary. (Global and Planetary Change. https://doi.org/10.1016/j.gloplacha.2020.103400)

Cretophengodidae, a new Cretaceous beetle family sheds light on the evolution of bioluminescence. (Proceedings of the Royal Society B. https://doi.org/10.1098/rspb.2020.2730)

Beetle or roach: systematic position of the enigmatic Umenocoleidae based on new material from Zhonggou Formation in Jiuquan, Northwest China, and a morphocladistic analysis. (Palaeoworld. https://doi.org/10.1016/j.palwor.2021.01.003)

Angiosperm pollinivory in a Cretaceous beetle. (Nature Plants. https://doi.org/10.1038/s41477-021-00893-2)

Revisiting the morphology and systematic placement of the enigmatic Cretaceous ommatid beetle Bukhkalius lindae (Coleoptera: Archostemata: Ommatidae). (Papéis Avulsos de Zoologia. https://doi.org/10.11606/1807-0205/2021.61.53)
New species of Paraodontomma from mid-Cretaceous Burmese amber with muscle tissue preservation (Coleoptera: Archostemata: Ommatidae). (Papéis Avulsos de Zoologia. https://doi.org/10.11606/1807-0205/2021.61.53)

Widespread mimicry and camouflage among mid-Cretaceous insects. (Gondwana Research. https://doi.org/10.1016/j.gr.2021.07.025)
Quantifying plant mimesis in fossil insects using deep learning. (Historical Biology. https://doi.org/10.1080/08912963.2021.1952199)

The mid-Miocene Zhangpu biota reveals an outstandingly rich rainforest biome in East Asia. (Science Advances. https://doi.org/10.1126/sciadv.abg0625)
