
Fig. 1 Some charophytes from the Pingyi Basin. A-C. Feistiella anluensis; D-I. Peckichara varians.
The Cretaceous-Paleogene boundary (K/Pg boundary) marks the beginning of the Cenozoic, and is one of the most important geological boundaries. The end-Cretaceous extinction is one of five mass extinction events, leading to the extinction of ammonites and belemnites in the sea. Also it is the end of dinosaur’s age. The K/Pg boundary is recognized as the boundary clay and the GSSP for the base of Danian stage is at the base of the clay. However, it is difficult to find the boundary clay in non-marine strata. Charophytes are usually very abundant in the non-marine strata near the K/Pg boundary, and the significant changes of charophyte flora can help to recognize the K/Pg boundary.
Recent investigation conducted by the Dr. LI Sha from “Modern terrestrial ecosystems origin and early evolution research team” at the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, cooperated with Carles Martín-Closas from the University of Barcelona, studied the change of charophyte floras from the Cretaceous–Paleogene transition in the Pingyi Basin, China and correlated it with European charophyte floras under the same age. The Cretaceous–Paleogene boundary was raised in the Pingyi Basin and the results were published on the Journal of Cretaceous Research.
Traditionally, the identification of charophytes in China is usually based on individual or a few gyrogonites but not the whole population. Such studies lack statistical measurements of the population and a uniform taxonomic standard, and do not consider the population characteristics and intraspecific variation. So it is difficult to make an intercontinental charopyhte biostratigraphic correlation and calibrate the K/Pg boundary in China. In study of the Pingyi Basin, Shandong Province, both traditions of identification from China and Europe were combined. For example, Porochara anluensis used in China was in fact belonged to the genus Feistiella according to observing the sections of basal plates. The intraspecific variations Peckichara varians was recognized based on population measurement and analysis. The abundant species Gobichara deserta around the Cretaceous–Paleogene boundary is in fact a younger synonym of Microchara cristata. (Fig. 1).
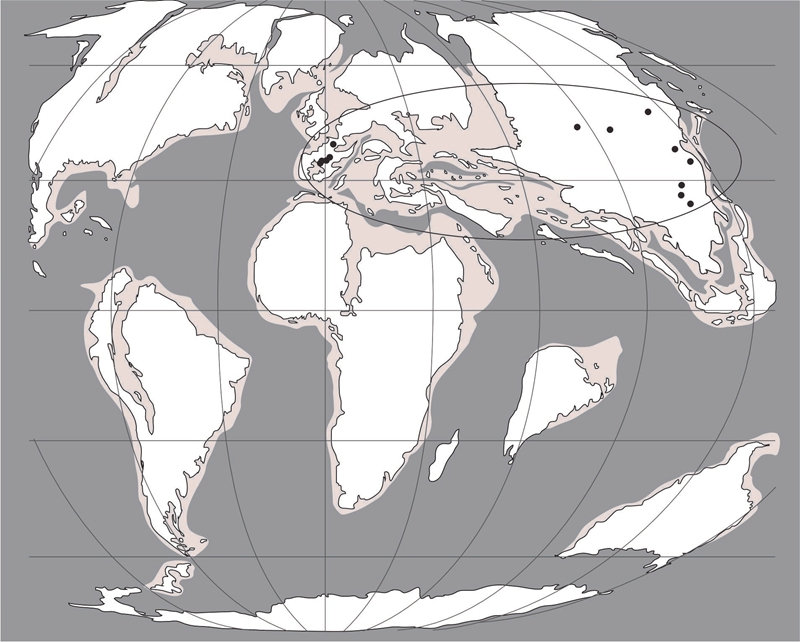
Fig. 2 Paleobiogeography of Microchara cristata in the Maastrichtian and early Paleocene, showing its Eurasian distribution. Paleogeography modified from Blakey (2006) with data plotted on a map at 65 Ma.
Based on a concensus of identification standard, a new biozonation is proposed which encompasses two biozones based on species with broad paleoecological requirements and a Eurasiatic distribution (Fig. 2, 3). European terrestrial Cretaceous–Paleogene boundary is mainly studied based on charophyte biozonations, the results can also be used in the research of Chinese terrestrial Cretaceous–Paleogene boundary. These are the Microchara cristata biozone starting in the latest Campanian and lasting at least until the earliest Danian and the Peckichara varians biozone encompassing the late Danian–earliest Eocene. The former can be correlated with biozones established by Galbrun et al. (1993) and Vicente et al. (2015) and calibrated to the GPTS, and the latter was correlated with Paleocene larger foraminifera in the Northern Pyrenees (Massieux et al., 1981). Therefore, the Cretaceous–Paleogene boundary from the Pingyi Basin was proposed to be in the first member (upper sub-member) of the Bianqiao Formation.
Reference: Li, Sha, Wang, Qifei, Zhang, Haichun, Lu, Huinan, Martín-Closas, Carles* (2016) Charophytes from the Cretaceous–Paleogene transition in the Pingyi Basin (Eastern China) and their Eurasian correlation.Cretaceous Research, 59: 179-200
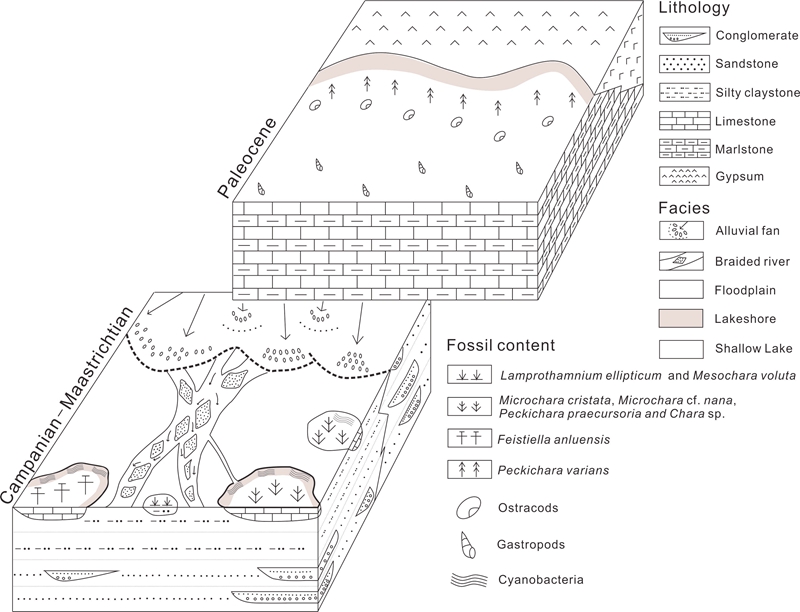
Fig. 3 Paleoenvironmental models summarizing the paleoecological distribution of charophyte floras in the Pingyi Basin (Gucheng and Bianqiao Formations) during the Maastrichtian and Paleocene.
